Xenasma praeteritum (H.S. Jacks.) Donk
Basidiomycota: Agaricomycotina: Agaricomycetes: Xenasmatales: Xenasmataceae: Xenasma ![]()
![]()
Introduction
The genus Xenasma consists of inconspicuous, whitish to bluish crusts. When fresh, they have a semitransparent, gelatinous appearance, but dry to a nearly invisible film. Unfortunately, these crusts can be as difficult to work with under the microscope as they are to see. The hyphae are small, gelatinized, and challenging to make out. The basidia and cystidia arise directly from this cryptic subiculum, which often appears embedded in the wood itself. While pleurobasidia are one of the defining features of Xenasma crusts, their hyphal origin is usually well hidden. But don't take my word for it — H. S. Jackson (1950) wrote the following about Xenasma:
The group as a whole is extremely difficult to study from dried material because of the gelatinous consistency when fresh and the vernicose ["brilliantly polished"] character when dry. Unless collections are dried quickly and were in active sporulating condition when collected, the basidia are difficult to make out. One is seldom able to view complete basidia or to trace them to the hyphae from which they originate.
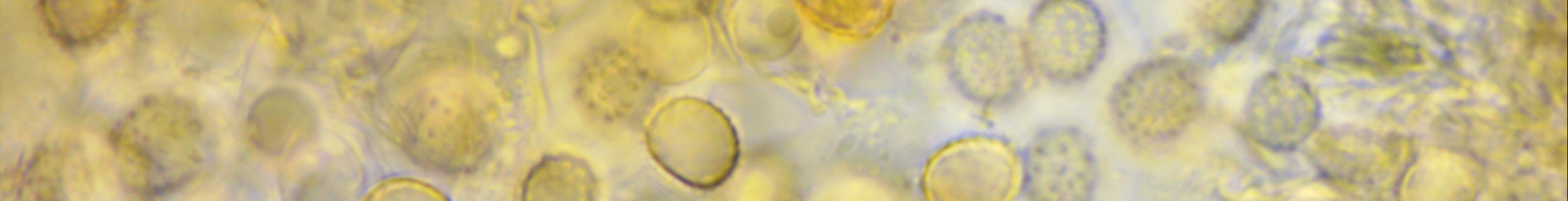
Fortunately, there is one defining character of Xenasma crusts that is easy enough to see: ornamented spores. However, in all but one species, the spore ornamentation dissolves in KOH, further complicating identification. Xenasma praeteritum is the only one whose spores do not do this, a feature that makes it perhaps the easiest species to recognize in the genus.
Description
Ecology: Presumably saprotrophic, on old, decorticated wood of deciduous trees, widespread across the temperate Northern Hemisphere.
Basidioma: Effused, smooth, ceraceous to gelatinous and somewhat transparent when fresh, drying to a subinvisible, pruinose film, margin undifferentiated and indeterminate, bluish.
Chemical reactions: NA
Spore print: NA
Hyphal system: Monomitic, individual hyphae very difficult to discern, clamped.
Basidia: Pleural (birooted), clavate to cylindrical, with two to four sterigmata (reported as three to five); length (23.8) 24–32.1 (34.9) µm, width (7.8) 8.0–9.4 (9.6) µm, x̄ = 28.1 ✕ 8.7 µm (n = 6 basidia).
Basidiospores: Broadly ellipsoid, somewhat thick-walled, ornamented (asperulate) in Melzer's reagent and KOH, inamyloid or weakly dextrinoid; length (6.7) 7.0–8.0 (9.2) µm, width (5.0) 5.5–6.6 (7.4) µm, x̄ = 7.5 ✕ 6.1, Q (1.1) 1.2–1.3 (1.4) (n = 30 basidiospores).
Sterile structures: Leptocystidia present, projecting well beyond the hymenium, thin-walled or somewhat thick-walled towards the base, at least 70 µm long and about 8 µm wide.
Sequences: ITS rDNA (OM009268).
Notes: All measurements taken in 5% KOH stained with phloxine. The microcharacters displayed by this specimen were all in all a good fit for Xenasma praeteritum as described by Jackson (1950). The basidiospores were the right size and the prominent spore ornamentation did not dissolve in KOH. However, the leptocystidia, which showed a thickening of the walls towards the base, and the number of sterigmata, of which I never observed more than four, were more reminiscent of Xenasma rimicola.
Specimens Analyzed
ACD0185, iNat33478376; 24 September 2019; Escher Co-op Woods, Washtenaw Co., MI, USA, 42.2949 -83.7244; leg. & det. Alden C. Dirks, ref. Bernicchia & Gorjón (2010), Jackson (1950); University of Michigan Fungarium MICH 352190.
References
Jackson, H. S. (1950). Studies of Canadian Thelephoraceae VI: The Peniophora rimicola group. Canadian Journal of Research, 28(5), 525–534.
Luo, K.-Y., & Zhao, C.-L. (2022). Morphology and multigene phylogeny reveal a new order and a new species of wood-inhabiting basidiomycete fungi (Agaricomycetes). Frontiers in Microbiology, 13(1966), 1–17.
Links

Ceraceous to gelatinous fresh basidioma.

Dried basidioma, visible in bright light.

The dried basidioma has a pruinose appearance under magnification due to the projecting cystidia.
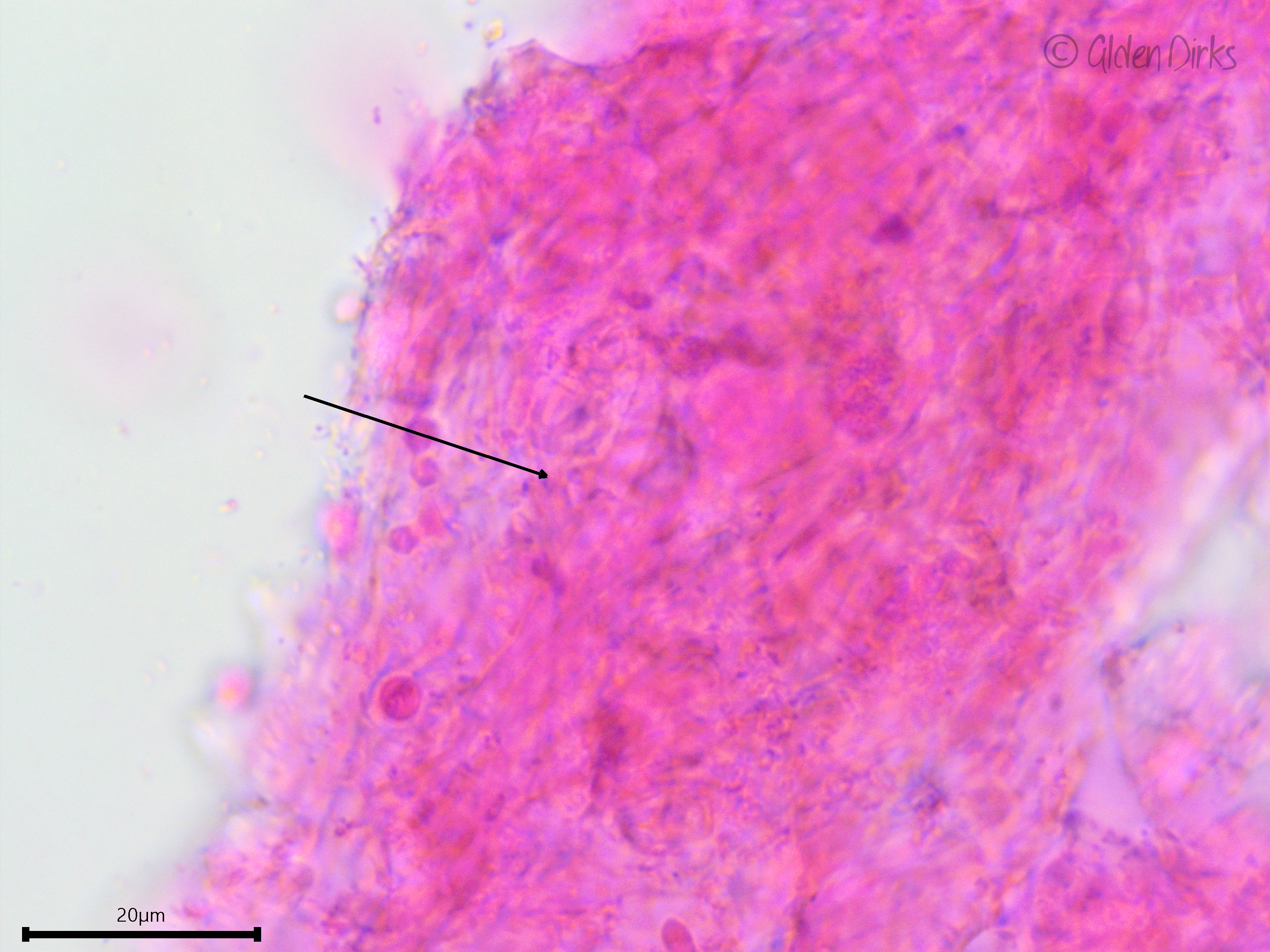
Hyphae are very difficult to discern. The arrow is pointing to a clamp connection.
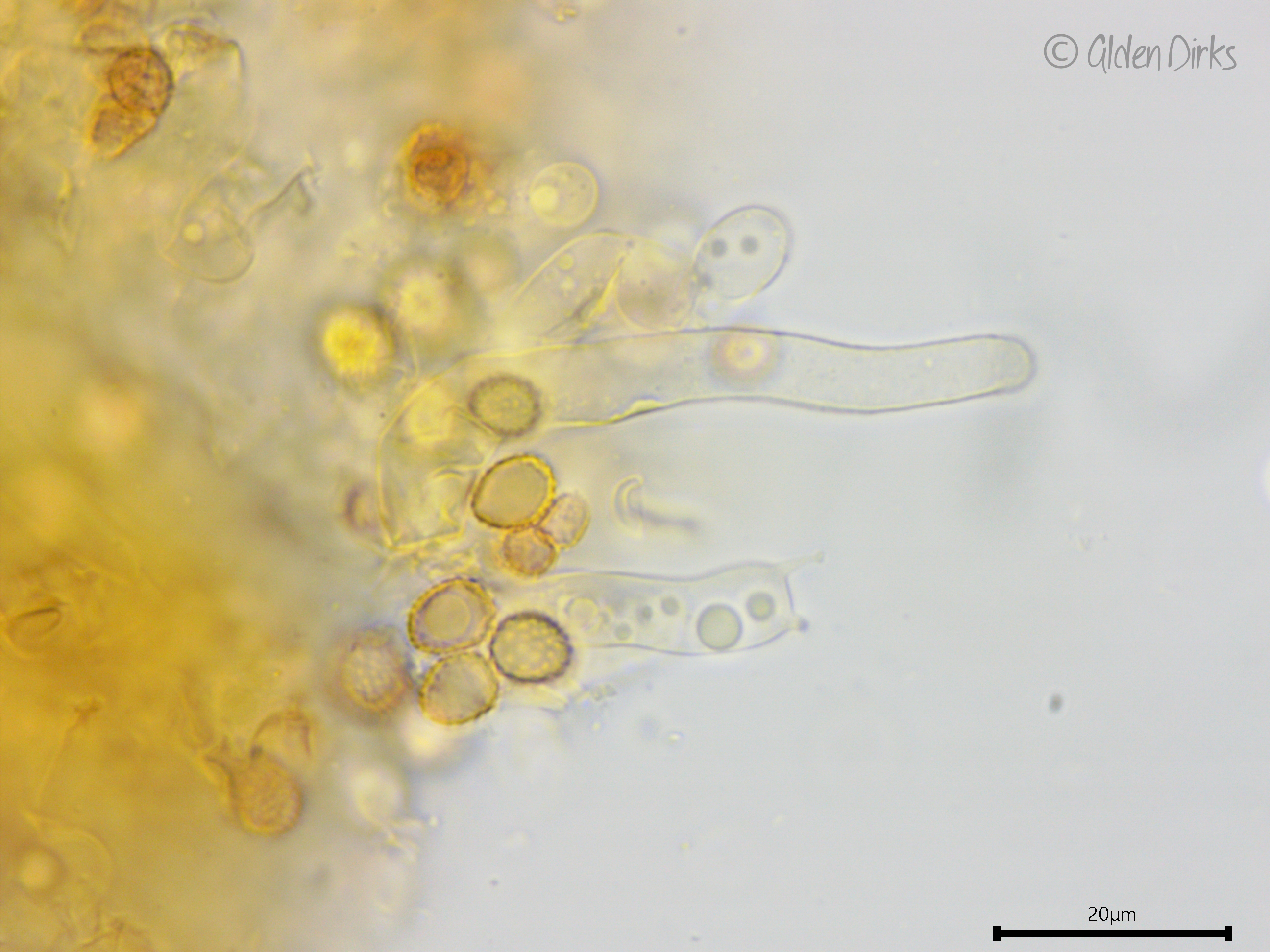
Developing basidium, weakly dextrinoid and ornamented basidiospores, and leptocystidium (Melzer's reagent).
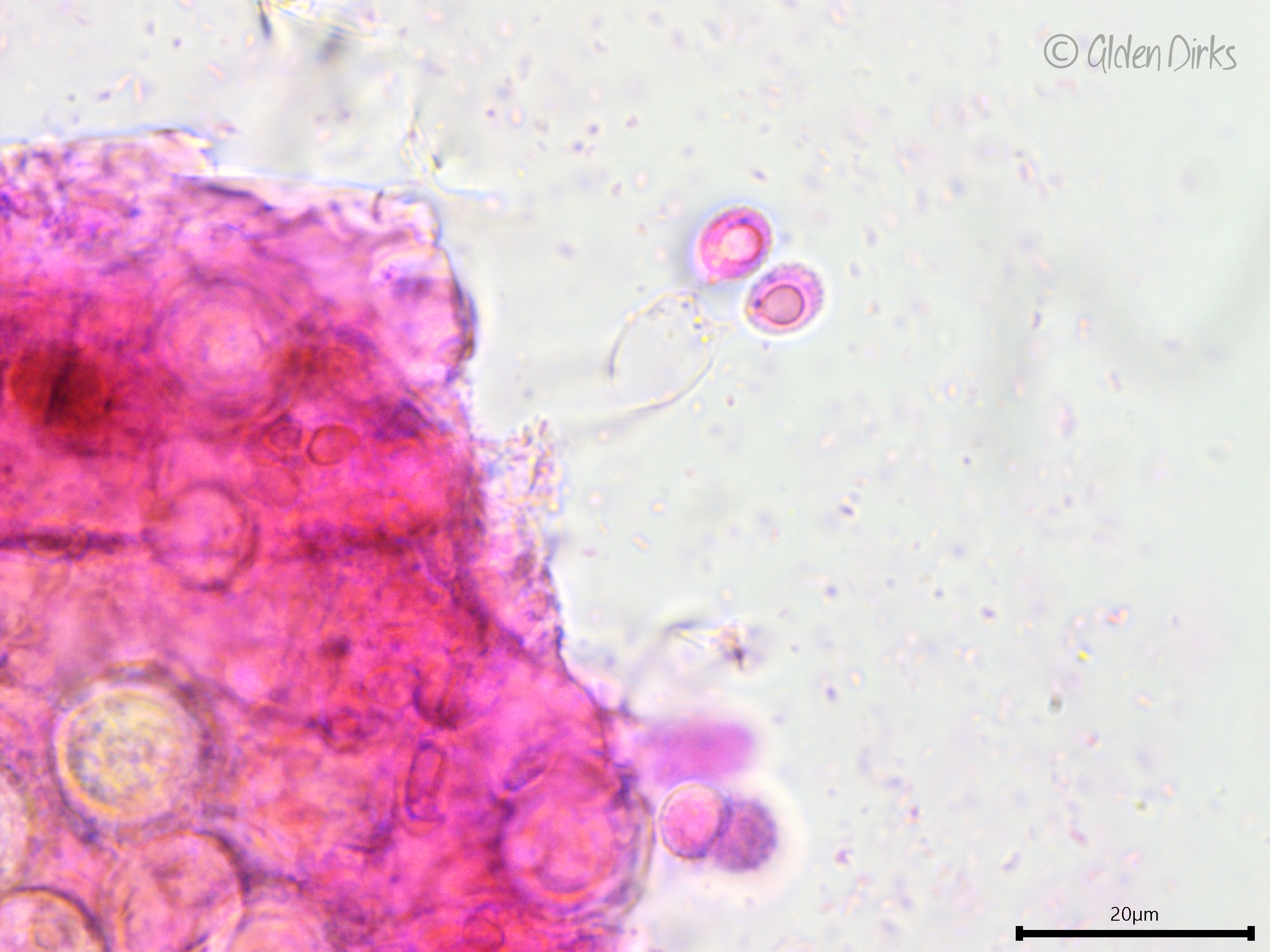
Basidium with two sterigmata.

Developing pleurobasidium.
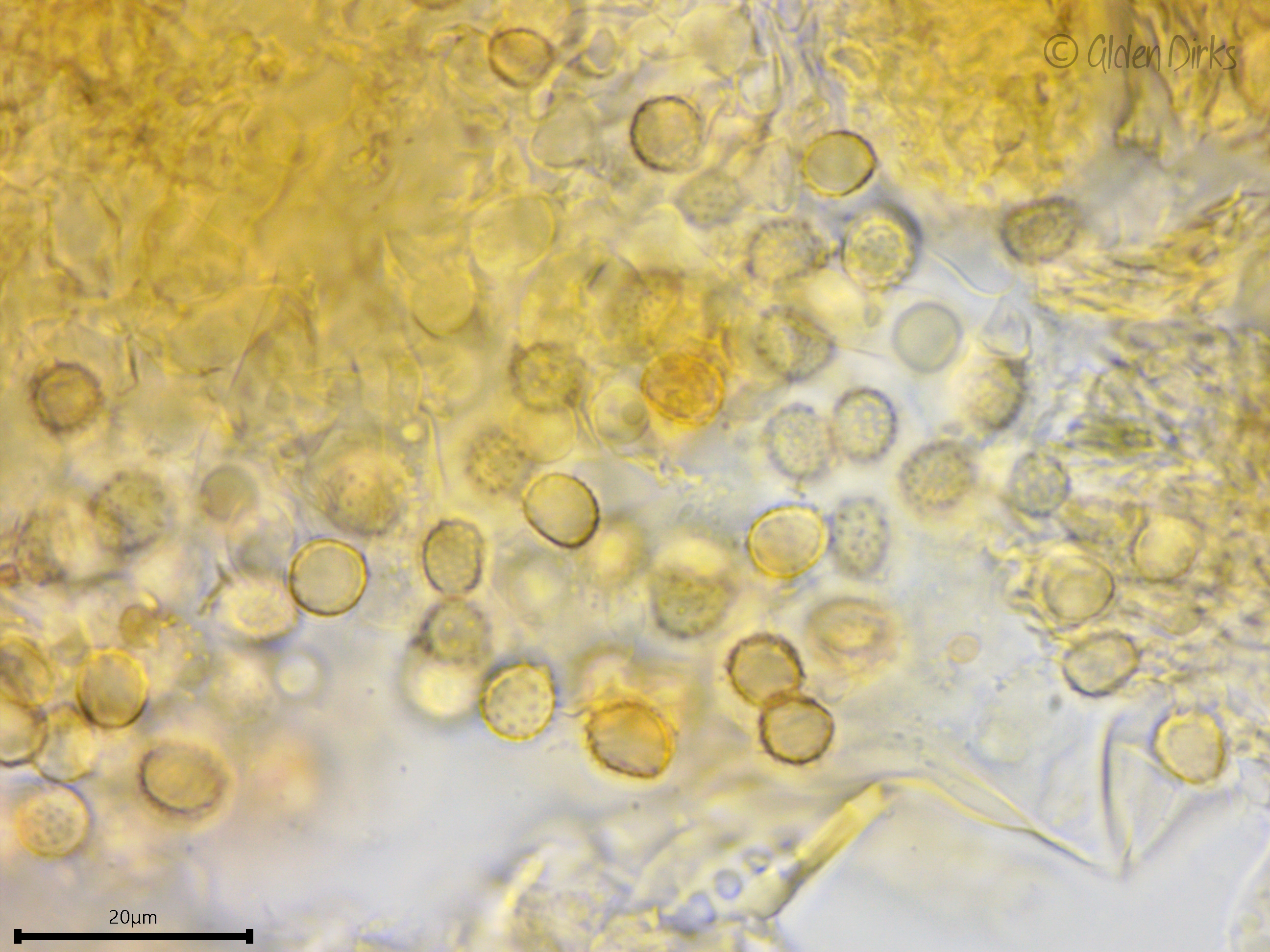
Ornamented basidiospores.
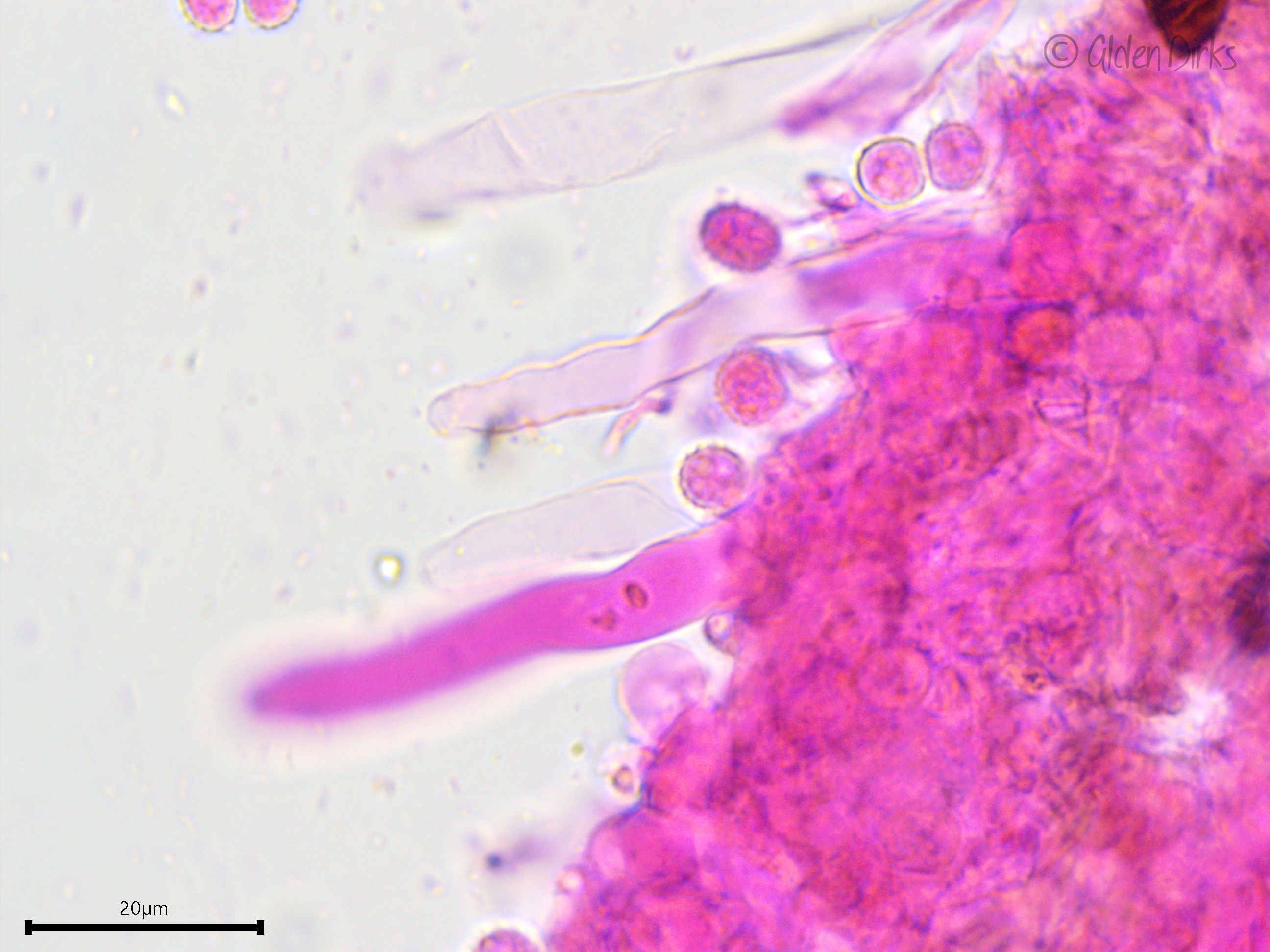
Leptocystidia.

Leptocystidium showing slight thickening of the cell wall towards the base.

Phylogenetic tree of Xenasmataceae.